Конкуренция среди водорослей и проблема выбора видов, перспективных
для санитарной марикультуры
О.Н. Селиванова
Промысловые водоросли российского побережья Северной
Пацифики (Восточной Камчатки и
Командорских островов) представлены, главным образом, бурыми
водорослями родов Alaria и
Laminaria. Их естественные запасы в наших северных водах еще не подорваны
промыслом, в отличие от более южных районов России (Приморье,
Сахалин). Поэтому на первый взгляд, в настоящее время нет
необходимости культивировать промысловые водоросли на Камчатке. Это
действительно так, если речь идет
о промышленной
марикультуре, т.е. об их пищевом
использовании. Но такая необходимость возникает для создания
санитарной марикультуры, т.е.
искусственного выращивания водорослей, активно поглощающих токсичные
вещества из морской воды, с целью их последующего удаления и
утилизации (например, сжигания или закапывания в почву).
Технологические приемы, оборудование и объекты
культивирования как для промышленной,
так и для санитарной
марикультуры, могут совпадать. Так, при
создании санитарной марикультуры могут
выращиваться водоросли, которые в общепринятом смысле являются
промысловыми (в нашем случае, это ламинариевые водоросли).
Различаются, очевидно, только цели и конечные результаты
культивирования: потребить или уничтожить культивируемый объект. Но
разработки, опробованные для одного из видов
марикультуры, могут быть впоследствии с
определенными модификациями использованы и для другого.
В настоящее время актуальна задача внедрения санитарной
марикультуры на Камчатке, которая вызвана
неблагополучной экологической обстановкой в Авачинской губе,
необходимостью очистки загрязненных вод этого водоема. Сочетание
биологических методов очистки, основанных на использовании донных водорослей и ряда морских беспозвоночных в
качестве фильтраторов, с техническими
средствами, очевидно, могло бы дать устойчивые положительные
результаты.
Идея
использования способности ряда бурых водорослей
к аккумуляции вредных веществ для очистки загрязненных вод закрытых
водоемов высказывалась нами в ряде предыдущих публикаций (Selivanova, 1996a, b, c; 1997; 1998; Селиванова, 1998а), в которых речь шла об использовании
водорослей из природных популяций. Дав предварительную оценку
поглотительным способностям бурых водорослей, произрастающих в зонах
Авачинской губы, подверженных долговременному воздействию
промышленного и бытового загрязнения (Селиванова, 1998а, Selivanova, 1998), мы встали перед необходимостью выбора наиболее подходящего
объекта для искусственного культивирования, поскольку природные запасы
водорослей необратимо сокращаются.
По данным атомно-адсорбционной
спектрофотометрии, среди
проанализированных 5 видов бурых водорослей из порядков
Laminariales и
Fucales, самой высокой способностью к аккумуляции вредных веществ (в
частности, тяжелых металлов) из загрязненной морской воды обладал
Agarum clathratum, значительно превосходя в этом отношении другие ламинариевые
водоросли, включая
Laminaria bongardiana
(Селиванова, 1998а).
Однако для решения задачи
внедрения санитарной марикультуры должны
быть учтены не только чисто “поглощающие” способности вида, но и его
природная численность, особенности роста и развития, а также
конкурентные отношения с другими видами. Так, например, в природных
условиях вышеупомянутый
A. clathratum уступает другим видам
ламинариевых по численности и по способности к конкуренции.
Проблемы конкурентных отношений между промысловыми водорослями
рассматривались в работах ряда российских исследователей (например,
Гайл, 1936;
Пржеменецкая, 1988) и большого числа
ученых из других стран Тихоокеанского региона (Японии, Китая, Кореи).
Однако мы можем использовать этот уже имеющийся опыт лишь в
незначительной степени, так как данные разработки относятся к видам из
более южных районов побережья Тихого океана. В наших районах
произрастают другие виды водорослей, к тому же мы имеем иные климатические условия. Поэтому нам необходимо
разработать свои подходы к решению проблем создания марикультуры на Камчатке. Мы
должны учесть биологические особенности произрастающих здесь видов
водорослей, определить виды-конкуренты и исключить их совместное
культивирование, а также выбрать те из них, которые обладают наиболее
широким спектром адаптаций к среде обитания.
В данной статье рассматриваются вопросы конкуренции среди ряда
водорослей, произрастающих у берегов Камчатки, их адаптивных
возможностей и жизненной стратегии. Помимо научного интереса эти
проблемы имеют и важное прикладное значение для выбора объектов
марикультуры.
Материалы и методы
Наше исследование основано, главным образом, на данных многолетних
полевых наблюдений за ростом, развитием и разрушением слоевищ
водорослей из зарослей келпа на прибрежных
мелководьях Камчатки и Командорских островов, постепенной заменой
одних видов и ассоциаций другими, и в меньшей степени, на
экспериментальных данных. В начале 90-х годов на о-ве Беринга
(Командорские острова) были выбраны 4 экспериментальных площадки для
наблюдений за ростом ламинарии (Laminaria bongardiana). Они располагались на скалистом рифе, защищенном небольшим мысом,
что предотвращало разрушительное воздействие океанских волн на
прибрежную растительность. На площадках проводили экспериментальное
выкашивание водорослей (на некоторых полное, на других частичное) и
наблюдение за последующим восстановлением и ростом ламинарии. Пластины
у той части растений, которые не выкашивались, были помечены
этикетками и перфорированы на разном уровне высоты от черешка. Прирост
пластины отмечался по увеличению расстояния между перфорациями через 1
и 11 месяцев после начала эксперимента (Иванюшина,
Жигадлова, 1994). Эксперименты проводили в
течение двух летних сезонов
1989 и 1990 гг. На полигоне №1 в 1989 г. было проведено полное
выкашивание всех ламинариевых водорослей крупнее 15 см; а в 1990 г. -
выборочное выкашивание 50 % макрофитов. На
полигоне №2 в первый сезон также проведено удаление
макрофитов на всей площади, а на следующий
год полигон был разбит на две части, на одной из которых повторно
выполнено тотальное выкашивание, а на другой водоросли оставлены
нетронутыми. На полигоне №3 два года подряд осуществляли
беспорядочное выкашивание 50 % ламинариевых
водорослей. И, наконец, на полигоне 4 работы проводили только в 1990
г. и они заключались в полном изъятии крупных слоевищ
макрофитов, а молодые растения были
оставлены нетронутыми (Ошурков, 2000). На всех полигонах растения
срезались при основании черешка ниже зоны роста во избежание
отрастания пластины в тот же сезон. Восстановление фитоценозов
оценивалось примерно в одни и те же сроки, чтобы исключить сезонную
вариабельность биомассы водорослей. Минимальная учетная площадь
составляла 0.25 м2. Оценку обилия крупных ламинариевых водорослей, взвешивание, подсчет
и измерение проводили на месте их сбора.
Замечание: при определении видовой принадлежности водорослей в
полевых условиях иногда появлялись ошибки. В частности, на
Командорских островах неверно были указаны L. gurjanovae и Hedophyllum sessile
(Иванюшина и др., 1991; Ошурков, 2000), но более тщательное
исследование в лабораторных условиях показало, что определенные так образцы, на самом деле, принадлежат
крайне вариабельному виду L. bongardiana.
Результаты и обсуждение
Исследования показали, что виды рода
Laminaria обладают достаточно высокой скоростью роста и являются очень
сильными и агрессивными конкурентами в борьбе за выживание. Среди них
нами отмечено несколько типов адаптаций к среде обитания, которые мы
подразделили на экологические, т.е. приспособления к различным
экотопам, различающимся по глубинам, типам
субстратов и т.п., морфологические (полиморфизм) и химические
(аллелопатия). Все эти типы адаптаций выражены в различной степени у
разных видов рода Laminaria (рис.
1). Так, L. bongardiana является
полиморфным видом, обладающим множеством жизненных форм, к тому же
этот вид имеет весьма широкий диапазон оптимальных глубин (от 0 до 30
м и более).
В зависимости от глубины произрастания изменяется и форма вида
(рис. 1) - от литоральной f. subsessilis (или
f.
bifurcata, которая, по-видимому, является возрастным вариантом первой) -
растения с коротким черешком и широкой, часто свернутой в виде
капюшона пластиной, до сублиторальной
f. taeniata -
c относительно длинным черешком и длинной узкой
ремневидной пластиной.
Типовая форма f. bongardiana является как бы
промежуточной между двумя перечисленными как по общему облику
растения, так и по глубине произрастания (рис. 1). Как видим,
L. bongardiana обладает сочетанием
морфологических и экологических адаптаций, и к тому же отличается
значительной скоростью роста. Такое сочетание свойств делает
L. bongardiana очень сильным
конкурентом по сравнению с другими видами, что вероятно, и
обеспечивает ее многочисленность, а точнее сказать, массовость у
берегов Камчатки и Командор.
Вторым по численности видом в этих районах является
L. dentigera, которая также отличается заметной скоростью роста и, кроме того,
имеет относительно большую продолжительность жизни.
L. dentigera нередко образует в
верхней сублиторали
монодоминантные сообщества, или чистые
заросли, состоящие только из растений этого вида.
Два вышеперечисленных вида ламинарий вступают в жесткую конкуренцию
за субстрат. По наблюдениям В.В.Ошуркова и
Е.А.Иванюшиной (Oshurkov and Ivanjushina, 1993) на шельфе о-ва Беринга после удаления L. dentigera с субстрата в результате промысла происходит ее замещение на более
короткоживущий вид
L. bongardiana. В экспериментах по
восстановлению сообществ ламинариевых через год после тотального
выкашивания наблюдалось уменьшение биомассы L. dentigera
при одновременном резком возрастании обилия
L. bongardiana. Иная картина наблюдалась после 50%-ного выкашивания: биомасса L. dentigera
через год восстанавливалась почти до прежнего уровня, а численность и
биомасса L. bongardiana существенно
уменьшалась (Ошурков, 2000). Можно предположить, что и природные
бидоминантные сообщества этих двух видов
ламинариевых находятся в
неустойчивом равновесии, при этом доля каждого из видов в
ценозе имеет обратную зависимость от доли
другого. При определенных обстоятельствах (оптимальная глубина,
освещенность, гидродинамика, предпочтительное
выедание вида-конкурента фитофагами и
т.п.) один из видов может приобретать неоспоримое преимущество, и
тогда формируется
монодоминантное сообщество. Чаще всего на
шельфе Командорских островов таковое образует L. dentigera на глубинах 6-12 м.
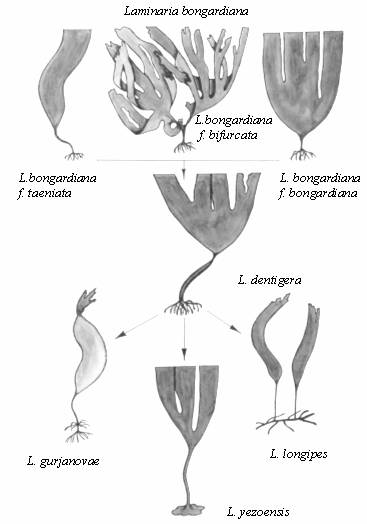
Рис. 1. Иерархия конкурентных способностей видов рода Laminaria
(уменьшение конкурентоспособности сверху вниз).
Аналогичные пары конкурирующих видов ламинарий, образующих сходные
по структуре и межвидовым взаимодействиям сообщества, выявлены и на
северном побережье Атлантики и Арктики: это L. digitata + L. longicruris (Атлантическое побережье
Канады) (Smith, 1986); L. hyperborea + L.saccharina
(северо-восточное побережье Атлантики) (Siversten, 1991) и L. digitata + L. saccharina
(Белое море) (Михайлова, 2000).
Остальные три вида ламинарий, произрастающих в наших водах,
L. gurjanovae, L. longipes, L.
yezoensis
- менее конкурентоспособны, хотя и у них имеется запас различных
вариантов адаптаций. В частности, можно отметить межвидовой
полиморфизм по типам органов прикрепления: пучок ризоидов у
L. gurjanovae, ризом у
L. longipes и диск у
L. yezoensis (рис. 1). Благодаря
этим морфологическим различиям, виды
дивергировали по разным экологическим
нишам: L. longipes стала
преобладающим видом в верхнем горизонте
сублиторали и нижней литорали,
L. yezoensis встречается на больших
глубинах, хотя оба вида произрастают на твердом грунте (скалистом или
каменистом). А вот
L. gurjanovae, благодаря
особенностям своих органов прикрепления, оказалась способной занимать,
в общем-то, несвойственные роду
Laminaria субстраты.
L. gurjanovae
растет не только на камнях и скалах, но и на рыхлых грунтах (песчаных,
илисто-песчаных, галечно-илистых и т. п.). У
L. gurjanovae, растущей в таких условиях у берегов Восточной Камчатки, мощно
развиваются ризоиды для обеспечения прочного закрепления растений на
рыхлом грунте. Пучок длинных разветвленных ризоидов напоминает хорошо
развитую мочковатую корневую систему высших растений. Иногда такие
пучки располагаются в несколько
ярусов, обеспечивая более надежное
прикрепление водорослей к грунту. Еще одной уникальной особенностью
L. gurjanovae является повышенная
хрупкость ее пластины в осенний период. Пластина этого растения не
обтрепывается в верхней части, как это происходит осенью у других
ламинарий, а обламывается выше черешка в 10-20 см от него. Очевидно,
оставшаяся на черешке часть пластины сохраняется до следующего года и,
по мере роста новой пластины, оказывается на ее верхушке. Возможно,
это способствует более быстрому отрастанию новой пластины за счет
остаточного фотосинтеза старой части, и таким образом, повышает шансы
растения в борьбе за выживание (Selivanova, 1999).
Итак, виды рода Laminaria, произрастающие у берегов Восточной Камчатки и Командор, образуют
нечто вроде иерархической лестницы по своим способностям к
конкуренции, верхнюю ступеньку которой занимает наиболее
активная в этом отношении L. bongardiana,
а в основании находится L. yezoensis.
Различные виды ламинарий распределились по
экотопам, различающимся как по типу
субстрата, так и по глубинам, что позволяет им избежать жесткой
конкуренции. На основе анализа вертикального распределения и
численности видов в верхней сублиторальной зоне Командорских островов
(от 0 до 10-12 м) нами выделены 3 водорослевые ассоциации с
доминантами Laminaria
longipes, L.
dentigera
и L. bongardiana.
В зоне ламинариевых водорослей на глубинах 1-3 м доминирующим видом
является
L. longipes (до 1600
экз/м2 при биомассе до 10.5 кг/м2).
Виды рода Alaria
часто выступают в качестве сопутствующих видов или
субдоминантов в фитоценозах ламинариевых и
обладают оппортунистическими свойствами по отношению к ламинариям. В
частности в поясе
L. longipes значительного обилия
может достигать A. angusta (до 2.4
кг/м2) (Селиванова, 1987).

Рис. 2. Виды рода Alaria, нередко
являющиеся субдоминантами в фитоценозах ламинариевых и выступающие
оппортунистами по отношению к ламинариям.
Сообщество Laminaria
dentigera+
Alaria spp. распространено на глубинах
от 1 до 6-12 м. Средняя биомасса его у Командорских островов
составляет 9.2 ± 1.5 кг/м2
(собственно ламинариевых 4.6±1.1 кг/м2) (Иванюшина и др.,
1991). Основным ценозообразующим видом
является L. dentigera, но иногда в бидоминантных сообществах
значительного обилия достигает и
L. bongardiana (до 4.1 кг/м2), сопутствующими видами выступают
L. yezoensis и
L. longipes. Доминантом второго порядка в данном
сообществе являются водоросли рода
Alaria, причем наблюдается вертикальная стратификация видов:
A. marginata (рис. 2)
распространена на глубинах 1-7 м, а
A. fistulosa (рис. 2) - на глубинах
от 5 до 15 м. Водоросли рода
Alaria являются постоянным компонентом
пояса ламинарий, но в некоторых участках распределены пятнисто.
Алярии могут достигать биомассы до 6
кг/м2.
В пределах ламинариевого пояса с доминированием собственно ламинарии
нередко в качестве субдоминантов 1-3
порядка выступают другие представители семейства:
Cymathere
triplicata,
Thalassiophyllum
clathrus,
Agarum
clathratum, при этом биомасса
T. clathrus может достигать 3.6
кг/м2. Иногда два последних вида могут приобретать доминирующую роль в
сообществах бентоса. Интересно отметить, что в естественных условиях
T. clathrus и
A. clathratum практически никогда
не наблюдаются совместно, хотя глубины их обычного произрастания
перекрываются (Ivanjushina, 1994; Oshurkov and Ivanjushina, 1994; Селиванова, 1998б). Мы полагаем, что морфологическая близость
этих двух видов ламинариевых водорослей и их сходные экологические
потребности являются причиной жесткой конкуренции, что приводит к
взаимному исключению друг друга и невозможности образования совместных
ассоциаций (рис. 3), что согласуется с правилом
Гаузе (Христофорова, 1999).
Бентосные сообщества на шельфе Командорских островов с
доминированием
L. dentigera, L.
dentigera
+ L. longipes или
L. dentigera +
Cymathere triplicata отличаются
обедненным видовым составом и ни один из организмов бентоса, помимо
ламинариевых, не достигает
сколь-нибудь значительной
биомассы.
Видовое богатство в ассоциациях с доминированием
T. clathrus, напротив, очень велико, количество видов
эпибентоса в них достигает 70. Аналогичную
с T. clathrus
ценозообразующую роль может выполнять
также A. clathratum, в ассоциациях
которого также наблюдается значительное видовое богатство организмов
бентоса. По мнению ряда авторов (Ivanjushina, 1994; Oshurkov
and
Ivanjushina, 1994), обедненный видовой
состав сообществ бентоса с монодоминантом
L. dentigera, а также
олигодоминантных сообществ
L. dentigera +
Cymathere triplicata (или
Alaria fistulosa) объясняется нестабильностью этих сообществ, связанной с
относительно короткой продолжительностью жизни доминирующих видов (1-4
года), тогда как T. clathrus имеет
в несколько раз большую продолжительность жизни (до 13-15
лет судя по числу годовых колец на срезе
черешка), что создает более стабильные условия для существования
морской биоты. Своеобразная морфология
этой водоросли, многочисленные складки пластины с фестончатыми
перфорациями и ее спиральные витки вокруг стволика, образующие
чашевидные углубления, служат убежищем для бентосных организмов и
благоприятствуют их поселению на слоевище. Кроме того, на глубинах,
где обычно произрастает
T. clathrus, пертурбации редки, и
таким образом, образуются относительно стабильные
микроэкотопы.
Как уже сказано, T. clathrus и
A. clathratum образуют пару
конкурирующих друг с другом видов. Несколько сложнее определить их
биологические отношения с другими видами ламинариевых водорослей.
Однако есть основания полагать, что
T. clathrus и
A. clathratum обладают пониженной
конкурентоспособностью по сравнению с ламинариями. Так, например, в
ряде экспериментов по тотальному выкашиванию ламинариевых водорослей
на следующий год в восстановленном сообществе появлялся
T.
сlathrus,
который отсутствовал в исходном фитоценозе. Логично объяснить
появление этой водоросли в сообществе снижением конкурентного пресса
со стороны ламинарий и, по-видимому, видов рода Alaria. А отдельные случаи, когда
T. clathrus или
A. clathratum все же вклиниваются в
группировки Laminaria sp.+ Alaria spp.
и даже приобретают в них доминирующее по биомассе значение, скорее
всего, можно объяснить выеданием их
конкурентов (т.е. ламинарий и алярий)
фитофагами.
В частности, на шельфе о-ва Беринга в тех сообществах L. bongardiana,
где в качестве одного из
субдоминантов выступал
T.сlathrus, непременным и наиболее обильным представителем зообентоса
являлись морские ежи Strogylocentrotus pallidus
(Ошурков, 2000). Но чаще всего
T. clathrus и
A. clathratum вытесняются
ламинариями на иные глубины. При этом для камчатского шельфа
характерно, что фитоценозы
T. clathrus (или
A. clathratum), как правило, отмечаются глубже пояса Laminaria bongardiana + Alaria spp. Это правило не всегда действует на Командорах, где наблюдаются
нарушения вертикальной стратификации растительных ассоциаций. T. clathrus и A. clathratum
могут подниматься там в литоральную зону, оказываясь, таким образом,
выше пояса ламинарий, т.е. в данном случае происходит инверсия
растительных поясов.
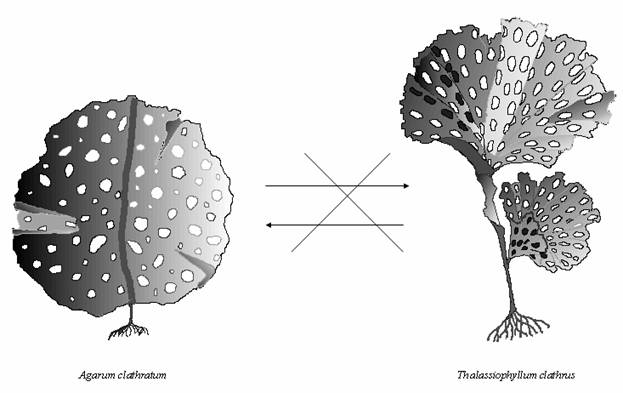
Рис. 3. Ламинариевые водоросли со сходной морфологией и
экологическими потребностями, которые являются конкурентами и не
образуют совместных сообществ.
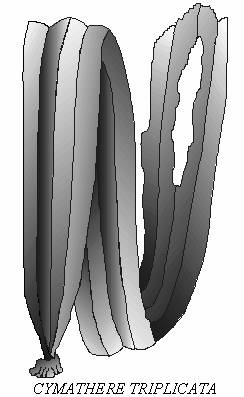
Рис. 4. Ламинариевая водоросль Cymathere triplicata, использующая химическое вещество с запахом огурцов в качестве
аллелопатического фактора.
В группу активных конкурентов, вероятно, следует отнести
Cymathere triplicata (рис. 4), хотя эта
водоросль и не столь многочисленна, как ламинарии, тем не менее, в
некоторых бухтах о-ва Медный она является доминирующим видом в
сообществах верхней сублиторали. Эта
водоросль обладает уникальной особенностью издавать запах свежих
огурцов в свежедобытом виде, чего не наблюдается более ни у одной
известной нам водоросли. Тот же запах встречается у других морских
организмов, в частности, у рыб семейства корюшковых
Osmeridae. Химическая природа и
биологическая роль этого запаха у
циматере пока не
выяснены. Но весьма возможно, что пахучее
вещество выполняет функцию
аллелопатического фактора, губительно
действующего на другие виды водорослей, и важного в конкурентной
борьбе, и таким образом, Cymathere является примером химической адаптации (Selivanova, 1999).
Таким же типом адаптации обладают корковые
кораллиновые водоросли. Их зона на шельфе
Командор имеет сравнимую значимость с сообществами ламинариевых. Эта
зона отличается значительным развитием корковых багрянок, образующих
слои известняка до 3-5 см толщиной. Наиболее обычным видом является
Clathromorphum nereostratum (рис. 5).
Значительно реже встречаются другие виды этого рода (C. circumscriptum
и C. compactum), а также виды родов
Lithothamnion и
Leptophytum. Исследованиями японских ученых (Masaki
et al., 1981;
Denboh et
al., 1998) было показано, что корковые
багрянки обладают способностью к активному вегетативному росту и
быстрому захвату свободных пространств субстрата, а также выделяют
аллелохимические вещества, обладающие
ингибирующими и репеллентными свойствами и
препятствующие росту и расселению ламинариевых водорослей. Из этого
следует, что кораллиновые могут быть
злостными сорняками при искусственном выращивании
ламинариевых в
марикультуре (рис. 5).
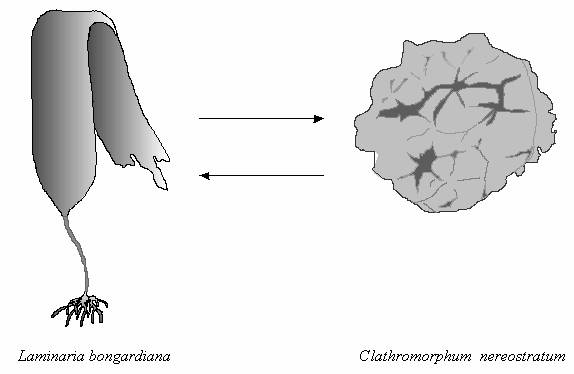
Рис. 5. Взаимное подавление ламинариевых и корковых
кораллиновых водорослей при колонизации
субстрата.
Однако многолетние данные по сукцессии бентосных сообществ на о-ве
Беринга свидетельствую о том, что победителем в конкурентной борьбе
между кораллиновыми и ламинариевыми в
итоге оказываются последние. Так, за десятилетний период с начала
1980-х до начала 1990-х годов обширные зоны у берегов о-ва Беринга на
глубинах 5-12 м, занимаемые прежде ассоциациями с доминированием C.
nereostratum, были замещены сообществами
ламинариевых.
Это могло произойти из-за целого ряда факторов 1) постоянного
разрушения корок кораллиновых под
истирающим воздействием подвижного гравия и камней в верхней
сублиторали при сильном прибое, 2)
малочисленности травоядных беспозвоночных (морских ежей) из-за
хищнического влияния калана 3) оппортунистической роли ламинариевых
по отношению к корковым
кораллиновым.
По нашему мнению, последний из перечисленных факторов наиболее значим.
Вероятно, в группу слабых в
конкурентном отношении представителей порядка Laminariales
из северной Пацифики следует включить Nereocystys luetkeana (сем. Lessoniaceae), Chorda filum (сем. Chordaceae) и Arthrothamnus bifidus (сем. Arthrothamnaceae) (рис. 6). В наших водах N. luetkeana
в настоящее время не произрастает, но он указывался ранее на
Командорских островах рядом исследователей (Гурьянова, 1935;
Зинова, 1940). Однако данные этих авторов
противоречат данным Е.А.
Кардаковой-Преженцовой (1938) и нашим
собственным многолетним наблюдениям на шельфе Командор (Селиванова,
1997). Мы встречали эту водоросль только в дрейфующем состоянии и в
выбросах, иногда в большом количестве, но никогда не обнаруживали ее
растущей на грунте. Современный ареал этой гигантской водоросли не
включает Командорские острова и ограничен на западе островами
Алеутской гряды Умнак и
Уналашка (Miller and Estes, 1989). Все же мы не можем
однозначно отрицать возможность произрастания N. luetkeana
на Командорских островах в прошлом и его последующее исчезновение в
результате сукцессии. По мнению
О.Г.Кусакина и М.Б.Ивановой (1995),
отсутствие этой водоросли в современной флоре островов связано с
флуктуациями обмена видами между Алеутскими островами и Командорами.
Явление резкой редукции численности и ареала другого
представителя гигантских
лессониевых водорослей Macrocystis integrifolia
наблюдается сейчас у берегов Северной Америки (личное сообщение д-ра
Гейл Хэнсен (Gayle Hansen), поэтому вполне вероятно, что такое же могло случиться и с N. luetkeana
в наших водах несколько ранее. Объяснения такой деградации гигантских
лессониевых водорослей могут быть самыми
различными, включая и неблагоприятные экологические условия, связанные
со все возрастающим антропогенным
воздействием. Однако, почему-то это не столь заметно на других
водорослях келпа. В таком случае причиной сокращения и исчезновения
лессониевых водорослей в ряде районов
Северной Пацифики логично было бы
предположить их пониженную конкурентоспособность по сравнению с
другими представителями порядка Laminariales. Примеры вытеснения N. luetkeana
под давлением более сильных конкурентов отмечены на северном побережье
США (штат Вашингтон) при восстановлении сообществ ламинариевых,
подорванных выеданием морского ежа Strongylocentrotus spp.
Реколонизация субстрата в этом случае
начиналась с нереоцистиса, но уже ко
второму, иногда к третьему году он замещался видами рода Laminaria (Paine and Vadas, 1969). У тихоокеанского побережья Канады (Британская Колумбия) при аналогичных обстоятельствах видовая
сукцессия приводила к третьему году к замещению ламинарии еще более активным конкурентом -
птеригофорой (Pace, 1981).
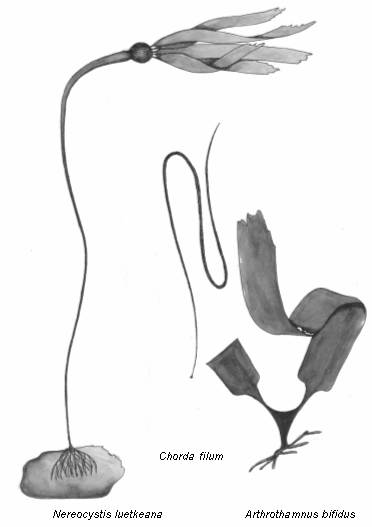
Рис. 6. Группа слабых в конкурентном отношении водорослей порядка
Laminariales.
Убедительными экспериментальными доказательствами слабой
конкурентоспособности двух других видов ламинариевых C. filum и A. bifidus,
мы пока не располагаем, однако их относительная малочисленность у
берегов Камчатки и ограниченность их ареалов свидетельствуют в пользу
такого предположения.
Проблемы конкурентных отношений среди основных промысловых
водорослей, к каковым принадлежат ламинариевые, представляют не просто
научный интерес, но имеют важное прикладное значение для создания
марикультуры. Объектами культивирования
следует избирать только те водоросли, которые смогут противостоять
натиску конкурентов.
Например, сколь бы ни была заманчива перспектива искусственного
выращивания гигантских
лессониевых водорослей в условиях,
близких к природным, (например, восстановление их якобы некогда
густых зарослей на Командорских островах), мы должны иметь в виду, что уже ко второму или третьему сезону
культивирования вместо запланированного урожая
N. luekeana, мы, скорее всего, получим плантации ламинарии.
Во всяком случае, такое явление отмечалось в природных условиях
(Paine and Vadas, 1969). При выращивании L. bongardiana
следует исключить попадание на субстрат спор L. dentigera,
иначе через несколько лет мы рискуем получить смешанное сообщество
двух видов ламинарии. Если это нас не устраивает, можно ограничить
срок культивирования 1-2 годами, и не дать возможности последнему виду
ламинарии вытеснить первый. Если же целью культивирования, наоборот,
является L. dentigera, нужно быть готовыми к тому, что первопоселенцем на искусственном
субстрате окажется L. bongardiana, а требуемую монокультуру нам удастся получить, возможно, лишь через
несколько лет.
Что касается санитарной
марикультуры, L. bongardiana,
несомненно, является более удобным объектом для культивирования, чем L. dentigera
(или какой-либо другой вид ламинарий), благодаря своей экологической
пластичности и устойчивости к повреждающему воздействию загрязнения,
что позволяет ей лучше справиться с задачей очистки морской воды, чем
другие виды ламинариевых. Кроме того, как было показано в
экспериментах (Ошурков, 2000), восстановление первоначального обилия L. bongardiana
после ее полного удаления с естественных субстратов происходит
довольно быстро, в течение 1-2 лет. Поэтому можно надеяться, что и при
удалении водорослей с искусственного субстрата (что является нашей
целью при внедрении санитарной
марикультуры), произойдет столь же быстрое
восстановление численности и биомассы L. bongardiana.
И хотя ее способности к аккумуляции вредных веществ значительно ниже,
чем, например, у A. clathratum, при
выборе перспективных видов для санитарной
марикультуры, следует отдать предпочтение
L. bongardiana, а не
A. clathratum, который более активен в поглощении
токсикантов, но обладает более низкой конкурентоспособностью.
Заключение
Данная работа является лишь попыткой выяснить конкурентные отношения
между основными промысловыми видами водорослей шельфа Восточной
Камчатки. Пока мы ограничились рассмотрением только вопросов
санитарной
марикультуры. Но в дальнейшем эти
исследования могут послужить основой и для будущего развития
промышленной марикультуры на Камчатке,
т.е. выращивания водорослей для пищевых целей, поскольку даже самые
богатые естественные ресурсы при их активной эксплуатации истощаются,
и возникает необходимость их восполнения. Поэтому
промышленная
марикультура является перспективной
отраслью развития любого прибрежного региона, где осваиваются
растительные запасы моря. Мы считаем, что наша работа должна быть
продолжена для того, чтобы выработать научно обоснованные рекомендации
для развития
марикультуры на Камчатке, что, в конечном
счете, поможет обеспечить долговременное и устойчивое развитие этого
региона.
Благодарности
Выражаю искреннюю признательность своим коллегам
Г.Г.Жигадловой и О.О.Новиковой за помощь в
подготовке и оформлении иллюстраций.
ЛИТЕРАТУРА
Гайл
Г. Ламинариевые водоросли дальневосточных морей // Вестник
ДВ Филиала АН СССР. 1936. Вып.19.
С.31-64.
Гурьянова Е.Ф. Командорские острова и их морская прибрежная фауна и
флора // Природа. 1935. №11. С.64-72.
Зинова
Е.С. Морские водоросли Командорских островов //
Тр. Тихоокеан.
Комитета. 1940. Вып.5. С.167-241.
Иванюшина Е.А., Жигадлова Г.Г. Биология
ламинарии Laminaria
bongradiana на литорали острова Беринга
(Командорские острова) // Биол. моря. 1994. Т.20. № 5. С. 374-380.
Иванюшина Е.А., Ржавский А.В., Селиванова
О.Н., Ошурков В.В. Структура и распределение
сообществ бентоса мелководий Командорских островов
// Природные ресурсы Командорских островов. Запасы, состояние, вопросы
охраны и использования. М.: Изд-во МГУ, 1991. С.155-170.
Кардакова-Преженцова
Е.А. Водоросли Командорских островов //
Изв. ТИНРО.
1938. Т.14. С.77-108.
Кусакин
О.Г., Иванова М.Б. Макробентос литоральных сообществ острова Медный
(Командорские острова) // Биол. моря. 1995. Т.21. №2. С.99-107.
Михайлова Т.А. Формирование ламинариевых фитоценозов на внесенном
каменистом субстрате в Белом море // Бот.
журн. 2000. Т.85. №9. С.88-103.
Ошурков В.В. Сукцессии и динамика
эпибентосных сообществ верхней
сублиторали бореальных вод. Владивосток:
Дальнаука, 2000. 206 с.
Пржеменецкая
(Макиенко) В.Ф.
Costaria costata (Huds.) Saund. (Phaeophyta,
Laminariales) в дальневосточных морях //
Комаровские чтения. 1988. Вып.35.
С.36-51.
Селиванова О.Н.
Макрофитобентос Командорских островов и
его особенности // Тез. докл.
науч.-практич.
конф. “Биол. ресурсы камчатского шельфа,
их рац.
использ. и охрана”.
Петропавловск-Камчатский, 1987. С.116-118.
Селиванова О.Н. Находка фрагментов бурой водоросли
Nereocystis luetkeana в Авачинской губе //
Биол. моря. 1997. Т.23. № 5. С.325-326.
Селиванова О.Н. Поглощение токсических элементов некоторыми бурыми
водорослями из загрязненных участков Авачинской губы // Сб. научных
статей по экологии и охране окружающей среды Авачинской бухты. 1998а.
Петропавловск–Камчатский: Изд-во
Госкомкамчатэкологии. С.39-45.
Селиванова О.Н. Макрофиты Командорских
островов (видовой состав, экология, распределение):
Автореф. дис.
… канд. биол. наук. Владивосток:
ДВГУ. 1998б. 25 с.
Христофорова
Н.К. Основы экологии. Владивосток:
Дальнаука, 1999.
516 с.
Denboh
T., Suzuki M., Mizuno Y., Ichimura T.
Supression of
Laminaria sporelings
by allelochemicals from coralline red
algae // Bot. Mar. 1997.
Vol. 40.
P.259-256.
Ivanjushina
E.A. Benthic communities of the
Masaki T., Fujita D., Akioka H.
Observations on the spore germination of
Laminaria japonica on
Lithophyllum yessoense (Rhodophyta, Corallinaceae) // Bull.
Fac. Fish.
Miller K.A., Estes
J.A. Western range extension for
Nereocystis
luetkeana in the
Oshurkov
V.V., Ivanjushina
E.A. Effect of experimental harvesting on
kelp regrowth and on the structure of the
shallow-water communities of
Oshurkov
V.V., Ivanjushina
E.A. The benthic associations of the
Pace D. Kelp community development in
Paine R.T.,
Vadas R.L. The
effects of grazing by sea urchins,
Strongylocentrotus spp., on benthic algal populations //
Limnol.
Oceanogr.
1969. Vol.14.
P.710-809.
Selivanova
O.N. Seaweeds as a possible ecological
factor of water purification and marine biota protection //
Abstr. PACON-96 International,
Selivanova
O.N. Adaptive strategy and possible
ecologic role of marine benthic algae // International
Sympos.
On Environmental restoration for
Selivanova
O.N. Benthic algae as a possible
ecological factor of water cleaning and protection of marine biota //
Abstr. II Russian-Japanese
Sympos.: Problems of Ecology and Nature
Management in
Selivanova
O.N. Participation of seaweeds in water
purification and marine biota protection // Recent Advances in Marine
Science and Technology (
Selivanova
O.N. Water cleaning ability of some kelp
algae // Abstr. PACON-98
International,
Selivanova
O.N. Competition between main commercial
algae of the
Siversten
K. Cleared areas and re-growth of kelp following harvesting operation
at Smola,
Smith B.D. Implications of population dynamics and interspecific competition for harvest management of the seaweed Laminaria // Mar. Ecol. Progr. Ser. 1986. Vol.33. P.7-18.