Питание щетинкочелюстных Parasagitta elegans в заливе Корфа
Берингова моря
В.В. Максименков
Щетинкочелюстные играют большую роль в морских экосистемах: они выедают в больших количествах веслоногих рачков (Sameoto, 1972; Sziper, 1978), иногда нападают на личинок рыб (Fraser, 1969), и сами являются пищей некоторых рыб (Андриевская, 1970).
Интерес к изучению питания щетинкочелюстных в заливе Корфа Берингова моря вызван тем, что здесь развиваются личинки промысловых видов рыб, таких как сельдь, минтай, камбалы, питающиеся мелким зоопланктоном. Щетинкочелюстные, вероятно, могут отрицательно влиять на их выживание в результате конкуренции за пищу или хищничества.
Parasagitta elegans - доминирующий вид щетинкочелюстных в заливе. В июне встречаются особи длиной 2-30 мм и крупнее, что приводит к возможности изучить также возрастные особенности питания сагитт.
Материал и методика
Щетинкочелюстные были собраны в заливе Корфа (1968-1984 гг.) икорными и
планктонными сетями в период учета численности личинок
корфо-карагинской сельди (июнь) на 45
станциях. Обилие сагитт оценено по уловам сети
Джеди (ячея - 0,11 мм) и выражено
в экз./м3.
Обработку фиксированного формалином материала проводили в лаборатории. У
парасагитт
измеряли длину тела и ширину головы. Затем животных вскрывали,
определяли таксономическую принадлежность жертв и измеряли ширину их
тела. Всего обработано 2334 экз. щетинкочелюстных, длина которых
изменялась в пределах 2-31 мм при
средней 12,1 мм. Суточные рационы рассчитаны
по методу Байкова (Baikov, 1935).
Результаты
Из 2334 экз. питалось всего 397 экз. (17%). Количество питающихся особей изменялось с длиной тела: 12,8% - у мелких сагитт; 32,4% - у животных длиной 10-11 мм; у взрослых сагитт доля питающихся была небольшой. Обычно в кишечнике встречалось по одному пищевому организму (91,8% случаев), реже - по два (5,4%), еще реже – по три и более (2,8%). Основу пищи щетинкочелюстных составляли веслоногие рачки (92,1% от количества), среди которых преобладали массовые виды зоопланктона Pseudocalanus minutus и Oithona similis (табл. 1). Личинки многощетинковых червей, усоногих и эвфаузиевых рачков, иглокожих, - также обильные в планктоне, в пище сагитт встречены редко. Отмечены случаи каннибализма. Икрой и личинками рыб щетинкочелюстные не питались. Самые мелкие особи использовали в пищу инфузорий из родов Parafavella и Tintinnopsis и науплиев веслоногих рачков; более крупные – в основном, копепод старших стадий развития и взрослых рачков. Доля микрозоопланктона в пище щетинкочелюстных снижается по мере увеличения размеров их тела (рис. 1).
Средний и максимальный размеры жертв являются функцией ширины головы хищника. В свою очередь, этот показатель зависит от длины тела парасагитт (см. формулы ниже). Таким образом, размеры потребляемых организмов увеличиваются по мере роста щетинкочелюстных.
Таблица 1.
Состав пищи (% от количества) мелких и
крупных P. elegans
|
Длина тела |
||
|
меньше 12 мм |
больше 12 мм |
|
|
Infusoria |
5,9 |
- |
|
Polychaeta
(larvae) |
0,5 |
|
|
Cladocera |
0,9 |
2,0 |
|
Ostracoda |
0,5 |
- |
|
Copepoda
(nauplii) |
16,4 |
7,0 |
|
Copepoda(copepodita et
adulta) |
67,7 |
80,4 |
|
Cirripedia
(nauplii) |
- |
1,0 |
|
Euphausiacea
(larvae) |
- |
0,5 |
|
Bivalvia
(veligri) |
1,8 |
1,0 |
|
Echinodermata
(larvae) |
- |
0,5 |
|
Chaetognatha
(juvenile) |
1,8 |
0,5 |
|
Appendicularia |
5,0 |
6,6 |
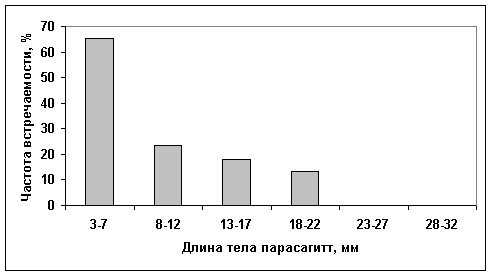
Рис. 1. Частота встречаемости мелких жертв (до 2,5 мм шириной) в пище P.elegans
различных размеров
Величины суточных рационов парасагитт рассчитаны по среднему количеству пищевых организмов в их кишечниках и продолжительности переваривания пищи (табл. 2). При этом были использованы полученные разными исследователями и построенные по собственным данным зависимости:
n * p
R = 24 * ────────── * 100 (Baikov, 1935),
N * P * D
P = 0,154 * W1,075 (Pearre, 1981),
W = 1.557 * h2,878 (Pearre, 1981),
h
= -0,030 + 0,991 * H – 0,610 * H2 (наши данные),
H = 0,051 * L0,967 (наши данные),
Log(P) = -10,032 + 3,245 * log(L) (Feigenbaum, 1982),
D = 10,240 * exp(-0,095 * t) (Kuhlmann, 1976-1977),
где R - суточный рацион, %; n/N - среднее количество съеденных одной парасагиттой организмов; W - масса сырого вещества парасагитты, мг; p, P - масса сухого вещества тела жертвы и хищника, соответственно, мг; h - ширина тела жертвы, мм; H - ширина головы парасагитты, мм; L - длина тела парасагитты, мм; D – продолжительность переваривания пищи, час; t - температура воды, градусы.
Суточные рационы по мере роста сагитт до модальных размеров увеличивались с 0.4 до 1.1 экз. или от .0004 до .0080 мг сухого вещества, а в дальнейшем снижались до 0.4 экз. или 0.0050 мг. Относительный рацион увеличивался с 6.0 до 9.9%, затем уменьшался до 1.0 %.
Располагая данными о численности сагитт в заливе Корфа и взяв величину суточного рациона для модальной размерной группы, можно ориентировочно оценить потребление мезопланктона щетинкочелюстными в разные годы (табл. 3). В зависимости от численности сагитт и температуры воды они выедали от 1 до 80 экз./куб. м в сутки.
Таблица 2. Исходные данные и оценка суточных рационов
парасагитты разных размеров
|
Длина тела, мм |
Среднее число жертв в кишечнике |
Масса жертвы, 10-3, мг |
Рацион, 10-4 мг сухого веса |
Относитель-ный
рацион, % |
|
|
4,5 |
0,14 |
8,0 |
0,4 |
3,6 |
6,0 |
|
5,5 |
0,19 |
13,5 |
0,6 |
9,0 |
8,1 |
|
6,5 |
0,24 |
20,4 |
0,7 |
16,1 |
8,5 |
|
7,5 |
0,31 |
28,4 |
0,9 |
29,7 |
9,9 |
|
8,5 |
0,24 |
37,2 |
0,7 |
31,5 |
6,8 |
|
9,5 |
0,32 |
46,3 |
1,0 |
57,0 |
8,8 |
|
10,5 |
0,36 |
55,3 |
1,1 |
75,9 |
8,3 |
|
11,5 |
0,38 |
64,0 |
1,1 |
80,0 |
6,6 |
|
12,5 |
0,21 |
71,9 |
0,6 |
54,6 |
3,6 |
|
13,5 |
0,25 |
78,7 |
0,8 |
80,0 |
3,9 |
|
14,5 |
0,18 |
84,2 |
0,5 |
54,0 |
2,1 |
|
15,5 |
0,20 |
88,2 |
0,6 |
67,8 |
2,1 |
|
16,5 |
0,14 |
90,5 |
0,4 |
46,4 |
1,2 |
|
17,5 |
0,15 |
91,1 |
0,4 |
46,8 |
1,0 |
Мы попытались оценить напряженность пищевых отношений среди личинок рыб и щетинкочелюстных по формуле Шорыгина:
![]() ,
,
где E –«сила конкуренции», a1i и a2i - суточные размеры потребления обоими видами данной группы организмов в весовых единицах на объем воды, b - биомасса данных кормовых организмов в тех же единицах на объем воды, d - степень пищевого сходства, g - поправка на географическое совпадение пастбищ (принята равной единице) (Руководство.., 1961).
Учитывая изменение удельного суточного рациона с длиной тела потребителей, было решено оценить величины рационов при средней длине и соответствующей массе хищников (табл. 4) и при предположении, что все личинки
Таблица 3. Потребление парасагиттами зоопланктона в разные годы
|
Год |
Температура воды, 0С |
Количество парасагитт, экз./м3 |
Потребление |
|
|
экз./м3 |
мг/м3 |
|||
|
1968 |
9,5 |
1,7 |
1 |
0,08 |
|
1969 |
8,4 |
6,9 |
4 |
0,31 |
|
1970 |
10,9 |
46,7 |
29 |
2,11 |
|
1973 |
9,3 |
5,9 |
4 |
0,26 |
|
1974 |
9,8 |
3,2 |
2 |
0,14 |
|
1975 |
8,8 |
21,5 |
13 |
0,97 |
|
1976 |
7,1 |
3,2 |
2 |
0,14 |
|
1977 |
8,3 |
133,0 |
80 |
6,02 |
|
1978 |
8,5 |
115,6 |
69 |
5,24 |
|
1981 |
10,9 |
8,1 |
5 |
0,37 |
|
1983 |
10,6 |
17,2 |
10 |
0,78 |
|
1984 |
9,5 |
84,6 |
51 |
3,83 |
Таблица 4.
Данные для оценки напряженности пищевых отношений в трофическом
комплексе (осредненные данные за 1976, 1977, 1980 гг.)
|
Потре-битель |
W, мг |
N, экз./м3 |
n/N |
W,
мг |
R,% |
P,
мг/сут.м3 |
Реаль-ный R, % |
Реаль-ное P, мг/сут.м3 |
|
Сельдь |
1,05 |
19,9500 |
2,98 |
0,010 |
8,3 |
1,7386 |
1,19 |
0,5945 |
|
Минтай |
0,53 |
0,0908 |
3,80 |
0,015 |
31,5 |
0,0152 |
4,94 |
0,0035 |
|
Песчанка |
2,65 |
0,0208 |
1,58 |
0,020 |
3,5 |
0,0019 |
0,32 |
0,0003 |
|
Бычки |
1,76 |
0,0205 |
5,09 |
0,030 |
25,4 |
0,0092 |
20,93 |
0,0010 |
|
Камбалы |
0,34 |
0,0358 |
2,38 |
0,008 |
16,4 |
0,0020 |
1,68 |
0,0009 |
|
Навага |
6,63 |
0,0078 |
6,23 |
0,050 |
13,8 |
0,0071 |
9,32 |
0,0005 |
|
Лисички |
2,24 |
0,0021 |
4,73 |
0,040 |
24,8 |
0,0012 |
15,69 |
1*10-4 |
|
Щетинко-челюст-ные |
0,70 |
13,8500 |
1,15 |
0,060 |
28,9 |
2,8019 |
5,03 |
0,1593 |
Примечания: W - средняя масса
потребителя, N - средняя численность
потребителя, n/N
- среднее число жертв в кишечнике потребителя,
w - средняя масса жертвы,
R -относительный (к массе потребителя)
суточный рацион, P -суточное
потребление.
рыб и щетинкочелюстные питаются.
Наибольшей численности среди прочих достигали личинки сельди,
численность личинок других рыб была на три порядка ниже, довольно
много встречалось щетинкочелюстных. Максимальный рацион оказался у
личинок минтая (31,5%), наименьший - у песчанки (3,5%), у личинок
сельди он не высок и в среднем составил 8,3%. Соответственно
численности, массе тела и величине рациона суточное потребление
оказалось максимальным у личинок сельди,
у щетинкочелюстных оно несколько ниже.
Ранее была рассчитана величина суточной продукции мелкого
зоопланктона, равная 5 мг/м3 (Максименков, Шагинян, 1991).
Суммарное потребление пищи рассматриваемыми видами составило бы 4,58
мг/ м3 или 91,5% от продукции. В действительности, реальные
средние рационы ниже, поскольку часть потребителей не питалась.
Соответственно, реальное суммарное потребление составило всего
15,2%.
Подставив данные таблицы 4 в формулу “силы конкуренции”, можно построить схему, показывающую напряженность пищевых отношений (рис. 2). Заметно, что щетинкочелюстные в заливе Корфа оказывают конкурентное влияние на личинок всех рыб, кроме сельди.
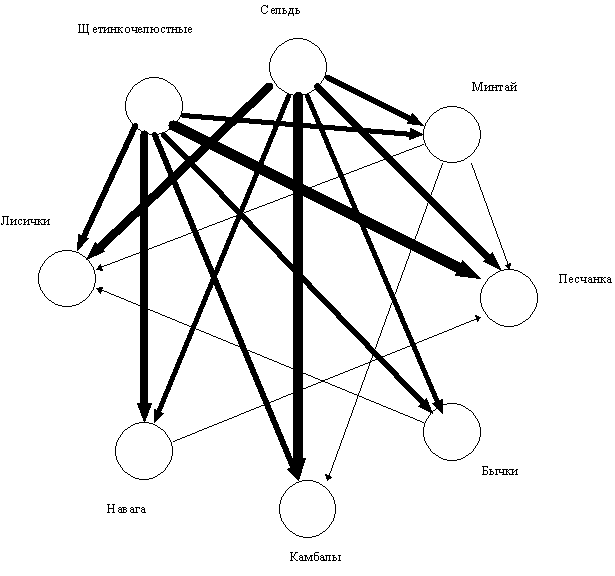
Рис. 2. Пищевые взаимоотношения личинок рыб и щетинкочелюстных в
Карагинском заливе. Толщина линии
пропорциональна силе влияния.
Обсуждение
Состав пищи сагитт и личинок, встречающихся в заливе рыб, в целом различен: если главной пищей личинок сельди, минтая, камбал являются велигеры двустворчатых моллюсков, яйца и науплии копепод (Максименков, 1984), то щетинкочелюстные питаются в основном, копеподитными стадиями веслоногих рачков, т.е. более крупными и подвижными организмами. Тем не менее, самые мелкие сагитты часто поедают организмы небольших размеров, в том числе инфузорий, науплиев копепод и даже личинок двустворчатых моллюсков, которые согласно мнению Миронова (1960), не могут быть удержаны ротовыми щетинками сагитт. Таким образом, при изучении питания и пищевых взаимоотношений личинок рыб в заливе Корфа необходимо обращать внимание на питание и численность мелких сагитт.
Полученные нами результаты, в общем, не противоречат выводам других исследователей. Так, большинство специалистов отмечает преимущественное питание щетинкочелюстных копеподами (Reeve, 1964; Pearre, 1973; Kuhlmann, 1976-77; Feigenbaum, 1982), низкий процент питающихся особей (Nagasawa, Marumo, 1972; Sziper, 1978; Feigenbaum, 1982), небольшое количество организмов в кишечнике (Миронов, 1960; Sziper, 1978).
По нашим данным, относительный рацион сагитт изменялся в пределах 1.0-9.9% в зависимости от размеров тела. Сходные величины получены другими исследователями (Дриц, 1981; Слабинский, 1982; Sziper, 1978; Feigenbaum, 1982).
Выводы
1. P. elegans питается, в основном, веслоногими рачками. По мере роста щетинкочелюстных увеличиваются размеры тела их жертв.
2. Суточные рационы по мере роста сагитт до модальных размеров увеличивались с 0.4 до 1.1 экз. или от 0.0004 до 0.0080 мг сухого вещества, а в дальнейшем снижались до 0.4 экз. или 0.0050 мг. Относительный рацион увеличивался с 6.0 до 9.9%, затем уменьшался до 1.0 %.
3. В зависимости от численности сагитт и температуры воды они выедали в разные годы от 1 до 80 экз./куб. м в сутки.
4. Щетинкочелюстные в заливе Корфа оказывают конкурентное влияние на личинок всех рыб, кроме сельди.
ЛИТЕРАТУРА
Андриевская Л.Д. Питание молоди тихоокеанских лососей в Охотском море
// Изв. ТИНРО.
1970. Т.78. С.105-115.
Дриц
А.В. Некоторые закономерности питания
Sagitta enflata // Океанология. 1981.
Т.21. №5. C.882-887.
Максименков В.В. Пищевые отношения личинок некоторых рыб в зал. Корфа
// Вопр. ихтиологии. 1984. Т.24. №6.
С.972-978.
Максименков В.В., Шагинян Э.Р. Продукция массовых видов
копепод и ее связь с температурой воды у
берегов Камчатки // Исследования биологии и динамики численности
промысловых рыб Камчатского шельфа. 1991.
Вып. 1. Ч. 2. Петропавловск-Камчатский:
КамчатНИРО. С.170-176.
Миронов Г.Н. Питание планктонных хищников //
Тр. Севастоп.
биол. станции. 1960. №13. С.78-88.
Руководство по изучению питания рыб в естественных условиях. М: АН
СССР. 1961. 262 с.
Слабинский
А.М. О питании массовых видов сагитт в заливе Петра Великого (Японское
море) // Изв.
ТИНРО. 1982. Т.106. С.80-83.
Baikov
A.D. How to estimate the daily food consumption of fish under natural
conditions // Trans. Amer. Fish.
Soc. 1935.
Vol.65. P.288-289.
Feigenbaum
D. Feeding by the chaetognath,
Sagitta elegans, at low temperatures in
Fraser J.H. Experimental feeding of some
medusae and
chaetognatha // J. Fish. Res.
Bd Can. 1969.
Vol.26. N. 7.
P.1743-1762.
Kuhlmann
D. Laboratory studies on the feeding behavior of the
chaetognaths
Sagitta cetosa Mueller and
S. elegans
Verril with special reference to fish eggs
and larvae as food organisms // Meer.
Rep. Mar. Res. 1976-77.
Vol. 25.
N.3-4. P.163-171.
Nagasawa
S., Marumo R. Feeding of pelagic
Chaetognatha
Sagitta nagae in
Pearre
S.Jr. Vertical migration and feeding in
Sagitta
elegans
Verril // Ecology.
1973. V. 54.
N. 2. P. 300-314.
Pearre
S.Jr. Feeding by
Chaetognatha: energy balance and
importance of various components of the diet of
Sagitta elegans // Mar. Ecol.
Prog.
Ser. 1981.
Vol.5. N.1.
P.45-54.
Reeve M.R. Feeding of zooplankton with
special reference to some experiments with
Sagitta // Nature.
1964. N201.
P.211-213.
Sameoto
D.D. Yearly respiration rate and estimated
energy budget for Sagitta elegans // J. Fish. Res.
Bd Can. 1972.
Vol.29. N7.
P.987-996.
Sziper G.P. Feeding rate of the chaetognath Sagitta enflata in nature // Estuar. Coast. Mar. Sci. 1978. Vol.7. N6. P.567-575.